
Abstract
Anthropogenic activities are increasingly encroaching into wildland areas, heightening interactions between human and carnivore communities. Area-based conservation measures, such as protected areas (PAs), employ different management strategies via land-use designations to mitigate anthropogenic pressures and reduce human-wildlife conflicts in shared landscapes. Here, we assessed carnivore diel activity and temporal activity overlap in and around El Triunfo Biosphere Reserve (REBITRI) in Chiapas, Mexico, along a land-use designation gradient. We deployed 33 camera traps along the gradient, leveraging the reserve’s core and buffer zones, and private lands surrounding the reserve. We calculated activity overlap between species to detect changes in interspecific competition and predator-prey interactions along the gradient. In total, we detected 14 carnivores in the core zones, 10 in the buffer zone, and 9 on private land across the 4777 trap-night survey. Significant shifts in single-species diel activity between the buffer zones and private land were detected for margay (Leopardus wiedii) and grey fox (Urocyon cinereoargenteus). Activity overlap was the highest in the buffer zone for all predator-prey pairs, and two competitor pairs, suggesting reduced diel niche partitioning in this land-use designation due to varied anthropogenic pressures. Our findings contribute to assessing PA efficacy and understanding carnivore activity in multiple-use landscapes where anthropogenic pressures are ubiquitous.
Similar content being viewed by others

Introduction
Protected areas (PAs) remain the primary conservation strategy for global carnivore protection to help maintain habitat connectivity, mitigate anthropogenic pressures, and support prey availability1,2,3,4,5. Large, continuous PAs benefit wide-ranging carnivores such as jaguars (Panthera onca) and pumas (Puma concolor)4,5. Proposals to increase the global PA coverage to 30% by 2030 in accordance with the Kunming-Montreal Global Biodiversity Framework are currently underway. Concurrent with PA expansion targets in the last two decades, the definition of PAs has broadened to include Other Effective Area-based Conservation Measures (OECMs). As described by the Convention on Biological Diversity, these PAs are governed by Indigenous peoples and local communities or other kinds of private landowners. OECMs are crucial for fostering justice and inclusion, ensuring diverse stakeholders are also involved in leading conservation efforts, creating more resilient PAs, and challenging fortress conservation approaches that were previously dominant6,7,8. As these inclusive area-based approaches become more eminent, conservation scientists and practitioners must now confront the formidable task of devising management strategies that promote coexistence between human and non-human communities in shared landscapes.
Shared landscapes are complex systems as wildlife are forced to adapt to increased levels of anthropogenic pressures. If left unmanaged, these pressures can progressively encroach upon the wildland areas, engendering heightened defaunation, habitat fragmentation, and human-wildlife conflicts9. Anthropogenic pressures are already rampant within the boundaries of traditionally designated PAs, raising global concern for biodiversity conservation10. Such disruptions that increase the probability of antagonistic interaction with humans can induce behavioral responses (i.e., movement, activity) in wildlife populations11,12. In the largest transboundary protected area in West Africa, livestock grazing shifted activity patterns in both ungulates and carnivores13. Other anthropogenic disturbances such as light pollution or bushmeat hunting are also known to affect wildlife by reducing their occupancy in disturbed areas and shifting foraging patterns14. Wide-ranging mammals are changing their spatial and temporal behaviors because of human-induced fear15,16. With the expanding human footprint in wildland areas projected to intensify by 2050, zoning of PAs through land-use designations has become a critical management strategy to protect biodiversity17,18.
In 2020, Mexico committed to increasing its PA coverage as part of the 30 × 30 international initiative19,20. Proper management and efficacy monitoring is an integral part of carnivore conservation in Mexican PAs and OECMs given that the decline in mammalian diversity in Mexico is largely driven by human-wildlife conflicts exacerbated by intensive land-use changes19,21. Mexican rural communities live in complex socio-ecological landscapes where increased interactions between humans and carnivores demand more concerted conservation efforts. A prime example can be found in Mexico’s tropical montane cloud forests, where key carnivore habitats and high levels of endemism coincides with agricultural areas dominated by small-scale coffee production22,23. This multiple-use nature of Mexican cloud forests poses challenges for conservation efforts that must maintain the economic viability of small-scale coffee plantations for local people while simultaneously promoting carnivore conservation.
Mexico’s montane cloud forests support a diverse carnivore guild, including large predators like the jaguar and puma, as well as mesocarnivores like the ocelot (Leopardus pardalis) and margay (Leopardus wiedii). Smaller carnivores often rely on temporal niche partitioning to minimize interspecific competition, with ocelots and margays adjusting their activity patterns where their ranges overlap24. Other mesocarnivores, such as the tayra (Eira barbara) and jaguarundi (Herpailurus yagouaroundi), exhibit diurnal behavior to avoid nocturnal competitors, while predator-prey interactions, like those between mesocarnivores and nine-banded armadillos (Dasypus novemcinctus) and common opossum (Didelphis marsupialis), also depend heavily on temporal niche partitioning25. Anthropogenic pressures, however, can disrupt these dynamics by driving shifts in diel activity as carnivores adjust to avoid antagonistic interactions or environmental stressors26,27. These shifts may reduce resource access by altering overlap with prey species and intensify intraguild competition among species with overlapping dietary niches, potentially triggering cascading effects on ecosystem stability24,28,29,30. Given that carnivores play an indispensable role in ecosystem functioning and hold high conservation importance in Mexico, balancing land-use changes and sustainable resource extraction with their conservation is imperative for promoting coexistence in shared landscapes25,31,32.
Here, we investigated how different land-use designations with varying management levels affect diel niche and activity overlap in a carnivore community. We conducted a camera trap study in El Triunfo Biosphere Reserve (REBITRI) in Chiapas, Mexico with cameras deployed in the core and buffer zones as well as outside of the reserve on private lands. Given the differences in management strategies and extraction activities, we utilized land-use designations as proxies for local habitat quality, prey availability, and human activity levels. We compared species-specific diel activity patterns and temporal overlap coefficients between focal species pairs across this land-use gradient. For single-species diel activity, we hypothesized that carnivores in core zones would exhibit the least variation in diel activity patterns compared to buffer zones and private lands, assuming increasing anthropogenic disturbance would drive greater deviations. On private lands, we anticipated further disruption of diel activity, reflecting pressure from unregulated extractive practices. We expected mesocarnivores such as ocelots to have greater shifts in diel activity in the buffer zone compared to the core zone. Since we expected larger carnivore detections to be low in private lands, the buffer zone is where mesocarnivores might experience the greatest combination of top-down pressures from both human activities and larger carnivores. For competing carnivore pairs, we hypothesized higher temporal activity overlap in private lands compared to core and buffer zones due to reduced niche partitioning under anthropogenic stress. We expected the level of activity overlap to be slightly higher in the buffer zones than in core zones for competing carnivores. Finally, for predator-prey interactions, we predicted reduced temporal overlap between diurnal carnivores (e.g., tayra, puma) and their prey species on buffer zones and private lands, assuming heightened human presence would drive diurnal carnivores to adjust diel activity. In contrast, we expected minimal changes in temporal overlap for nocturnal carnivores (e.g., ocelot, margay) and their prey on buffer zones and private lands, assuming human activity remains primarily diurnal. Our findings contribute to assessing PA efficacy and understanding carnivore response to anthropogenic pressures in shared landscapes.
Methods
Study area
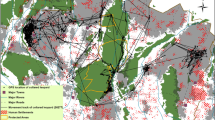
El Triunfo Biosphere Reserve is an IUCN Category VI protected area, spanning 1192 km2 in the state of Chiapas, Mexico (Fig. 1A). It contains the most extensive continuous evergreen cloud forest in the southern part of the country33, and is home to a diverse mammalian carnivore community, including jaguars, pumas, and ocelots. The reserve is well-known for its high bird biodiversity as well as high diversity of epiphytes and orchid species34. There are four main climate types in the reserve: subhumid tropical, humid subtropical, humid tropical, and humid temperate with the dry season extending from November to April34. The Sierra Madre de Chiapas (4220 m above sea level) bisects the reserve, generating microclimates from the highlands to the lowlands that promote high levels of endemism. Two other types of PAs, Protected Area of Natural Resources, La Frailescana and Area Subject to Ecological Conservation, Cordon Pico El Loro-Paxtal, border the northern and southern part of the reserve, respectively. The reserve has two distinct management levels: five core zones (21.9% of reserve area) and a buffer zone (78.1%). The reserve is a multi-use protected area, with the presence of over 40 rural communities mostly concentrated inside the buffer zone. Agricultural practices (i.e., small-scale subsistence and small-scale commercial agriculture) and cattle ranching are permitted in the buffer zone but are restricted in the core zones. A mix of rural communities and property for large-scale commercial agriculture are found in the surrounding areas, outside reserve boundaries.

El Triunfo Biosphere Reserve study area. (A) Camera trap survey design, showing four circular sampling units and camera stations established. (B) A land-use gradient was established by placing camera traps in the core zones, the buffer zone, and on private land outside the reserve.
Camera trap survey
We conducted a systematic camera survey in REBITRI from May to November 2023. Unbaited, Reconyx camera traps (Reconyx, WI, USA) were placed in the core zones, buffer zones, and on private lands outside the protected area with no enforced conservation-based management (Fig. 1B). The reserve was systematically divided into four circular sampling units, each approximately 22 km in diameter, capturing all three land-use designations. Within each unit, three cameras were deployed per land-use designation for a total of nine cameras. Due to constraints on access, we successfully deployed 33 of 36 planned camera trap stations (Core zones: n = 12, Buffer zones: n = 11, Private lands: n = 10). Camera traps were placed at ground-level (30–60 cm) near game trails to maximize carnivore detection probability35, and at least 1 km from each other to avoid detection overlap bias36. We programmed cameras to high sensitivity, with 1-second lapse between three images and a video per trigger, and a 15-s quiet period between triggers. Following the conclusion of the survey period, camera trap images were sorted into species-specific categories employing established identification guides and camera trap sorting software, Timelapse 2.0 image analyzer37. Subsequently, sorted images underwent a comprehensive review and validation process conducted by an independent researcher from the Applied Wildlife Ecology Lab not associated with the initial sorting procedure. To ensure accuracy and consistency, consecutive images of the same species captured within a 30-minute period at a specific camera trap ___location were filtered as a singular detection event using the camtrapR package in R (version 4.1.0)38. The activity patterns and their overlap were quantified for species displaying ten or more independent detections in at least two of the land-use designations.
Single-species diel activity
Time stamps for each independent detection event were extracted from the image metadata. Using time of day as a circular variable, diel activity patterns were constructed based on the distribution of captures across the hours of the day for carnivores with sufficient independent detections. Time ranges for light-dark cycles were defined for nocturnal (19:00--04:00), crepuscular (04:00–05:00, 18:00–19:00), diurnal (05:00–18:00) and cathemeral species in accordance with the region’s daylight cycle. We classified species’ diel activity patterns quantitatively using a time-bin approach39,40. Cathemeral activity was defined as detections with ≥ 20% activity during both nocturnal and diurnal time ranges. For each species and land-use designation, we calculated the proportion of detections in each diel period. Species were classified as crepuscular, diurnal, or nocturnal if ≥ 50% of detections occurred within their respective time ranges. Species were classified as cathemeral if activity was distributed more evenly, with ≥ 20% of detections occurring during non-peak periods. Diel activity was estimated using non-parametric kernel density estimation methods available in the ‘activity’ package in R (version 4.1.0)41. We used the compareCkern() function to compare activity estimations over time of day through a permutation-based approach. We conducted 10,000 permutations to generate the null distribution and assessed the significance of the observed differences in diel activity patterns between land-use designation pairs (i.e., core vs. buffer, core vs. private, etc.).
Temporal activity overlap for species pairs
We also compared activity overlap among species pairs that reflected both predator-prey and competitor dynamics across land-use designations. Using the ‘overlap’ package42 in Program R, we applied non-parametric kernel density estimation methods to generate species temporal activity overlap graphs41. We then quantified the overlap between species’ activity curves using the overlap coefficient “Dhat1” (∆1) using a smoothing parameter of 0.8 given all species analyzed had ≥ 75 independent detections41. We performed a Mardia–Watson–Wheeler (MWW) test to determine differences in the angular means between samples of circular data for species pairs using the ‘circular’ package in Program R. We also estimated confidence intervals around species pair activity overlap using 10,000 bootstrap iterations. If two species have a higher temporal activity overlap, we expect a higher overlap coefficient (∆1 ≥ 0.5), a lower Watson-wheeler statistic (W ≤ 6.0), and a non-significant p-value.
Results
Our total sampling effort spanned 4777 trap nights (1632 in core zones, 1630 in the buffer zone, 1515 on private lands) from May to November 2023. The occurrence of human detections did not follow our expectation and were highest in the buffer zone (n = 155), followed by the core areas (n = 134), and private lands (n = 58). Mule detections were limited to the buffer zone (n = 36), and domestic dogs were detected in the buffer zone and on private land (n = 19, n = 32, respectively). Per expectations, no domestic animals were detected in the core of the reserve. We observed a diverse carnivore guild with 14 species, all of which were detected in the core zone while 10 and 9 species were detected in the buffer zone and on private lands outside the reserve, respectively (Table 1). Tayra and puma were the species with the highest number of detections in the core zones (n = 29); the grey fox (Urocyon cinereoargenteus) presented the highest number of detections in the buffer zone (n = 90); and white-nosed coati (Nasua narica) on private lands (n = 65). Hooded skunk (Mephitis macroura), jaguarundi, long-tailed Weasel (Mustela frenata), cacomistle (Bassariscus sumichrasti), neotropical otter (Lontra longicaudis), jaguar, spotted skunk (Spilogale angustifrons), and raccoon (Procyon lotor) detections were excluded from the single-species activity and activity overlap analyses because they did not have at least 10 independent detections in at least two land-use designations.
Single-species diel activity shifts
We assessed differences in activity patterns throughout the time of day for single-species across the land-use gradient, expecting differences to be most significant when comparing activity in the core zones with activity in the buffer zone or on private land. Our proportional classification of diel period for each species and land-use designation revealed limited shifts in light-dark cycles (i.e., nocturnal or diurnal) among the carnivore species, with carnivores predominantly remaining within their respective diel niches (Fig. 2). Puma were largely cathemeral with an even percentage of nocturnal and diurnal detections across all land-use designations, and the highest levels of activity peaking in the early morning for both core and buffer zones. Tayra and coati exhibited minor shifts in their light-dark cycles, staying predominantly diurnal across all land-use designations with a high percentage of detections (> 85%) occurring during the diurnal time range. Margay displayed cathemeral activity in core zones and private lands, while the buffer zone had 70% of detections occurring during the nocturnal time range. Grey fox shifted from exhibiting cathemeral activity in the buffer zone to having a higher percentage of diurnal detections on private lands (75%) and core zones (100%), which was partially consistent with our hypothesis. Ocelot shifted from exhibiting cathemeral activity in both core zones and private lands, with unexpected peaks of activity during the daytime, to exhibiting higher percentage of nocturnal detections (70%) in the buffer zone. Our comparison of activity using a permutation-based approach revealed significant differences in diel activity throughout the time of day between land-use designations for only margay (core vs. buffer: p = 0.021) and grey fox (buffer vs. private: p < 0.001). Ocelot exhibited minor differences between core and buffer zones, though they were not statistically significant (p = 0.095; Table 2).

Kernel density estimation plots of diel activity for six Mexican carnivore species derived from a 4777 trap-night camera survey conducted in El Triunfo Biosphere Reserve. Top row: tayra, ocelot, gray fox; Bottom row: puma, margay, coati.
Species pairs activity overlap
We assessed temporal activity overlap analysis among species pairs to explore changes in predator-prey and competitor interactions along the land-use gradient (Table 3, Figure S1). Margay and ocelot showed no significant differences in overlap activityacross all three land-use designations (core: W = 1.98, p = 0.37; buffer: W = 2.64, p = 0.2; private: W = 3.43, p = 0.18). However, contrary to our expectations, there was higher activity overlap between this pair in the core area (core: ∆1 = 0.73, buffer: ∆1 = 0.69; private: ∆1 = 0.57). Conversely, another competitor pair, margay and tayra exhibited significantly different activity overlap across all land-use designations (core: W = 39.5, p < 0.001; buffer: W = 11.88, p = 0.003; private: W = 18.34, p < 0.001). Partially consistent with our hypothesis, activity overlap for margay and tayra was higher in the buffer zone (∆1 = 0.35) and private lands (∆1 = 0.25) compared to the core zone (∆1 = 0.12); although, we expected to see higher activity overlap in the private lands. Finally, we evaluated how activity overlap varied between puma, a large carnivore, and ocelot, a mesocarnivore. We found that activity between this pair was not significantly different in the core zones (W = 2.08, p = 0.35) but was marginally different in the buffer zones (W = 5.21, p = 0.07). Consistent with expectations, the overlap coefficients revealed that activity between puma and ocelot was higher in the buffer zones (∆1 = 0.67) compared to the core zones (∆1 = 0.59).
For the activity overlap analysis for predator-prey interactions (Figure S1), we compared the ocelot and nine-banded armadillo. Unexpectedly, activity levels were significantly different in the core zone (W = 10.03, p = 0.007) followed by a marginal significant difference on private lands (W = 5.95, p = 0.051) and no significant difference in the buffer zones (W = 2.7, p = 0.26). Contrary to our hypothesis for predator-prey species, core zones had the lowest overlap coefficient (∆1 = 0.43) with private lands trailing close behind (∆1 = 0.52), compared to buffer zones (∆1 = 0.72), which had a high overlap coefficient. We also analyzed the overlap activity of margay and the common opossum as predator-prey pairs across the land-use gradient. Activity levels for margay and common opossum were significantly different in core zones (W = 11.73, p = 0.003) and private lands (W = 9.80, p = 0.007), while no significant difference was found for buffer zones (W = 3.9, p = 0.14). Once again contrary to our expectations, the buffer zone had the highest overlap coefficient (∆1 = 0.68), while core zones and private land both had similar, lower overlap coefficients (∆1 = 0.63, ∆1 = 0.53, respectively). Mirroring our findings above, pairing the ocelot and common opossum also yielded different activity levels in the core zones (W = 17.45, p < 0.001) and private lands (W = 8.41, p = 0.015), while buffer zones had similar activity levels (W = 2.46, p = 0.29). Core zones and private lands had lower overlap coefficients (∆1 = 0.45, ∆1 = 0.55, respectively), but in the buffer zones, the overlap coefficient was much higher (∆1 = 0.80).
Discussion
Wildlife diel activity patterns are crucial for understanding species interactions and niche dynamics as anthropogenic pressures transform landscapes43,44. Our study sought to determine whether species-specific diel activity and temporal overlap shift across a land-use gradient encompassing core zones, buffer zones, and private lands. Our findings from the REBITRI area indicated varied diel activity and temporal overlap across different land-use designations. Surprisingly, out of the seven carnivores analyzed, only the margay and grey fox showed significant differences in diel activity across zones, with the most pronounced shifts occurring in the buffer zone. Additionally, we found that the buffer zones had the highest temporal overlap in activity for all predator-prey pairs and two competitor pairs. Promoting coexistence between carnivores and humans requires a comprehensive understanding of carnivore ecology and behavior, along with inclusive conservation approaches that align conservation goals with improvements in human livelihoods. At a global level, these issues are expected to increase as shared landscapes where human and non-human communities are present become declared as PAs to meet biodiversity targets.
The absence of large carnivores results in a lack of top-down regulation that often releases mesocarnivores to widen their niche space45,46. Given variations in a species’ sensitivity to disturbance, the land-use designation used in our study likely serves as a proxy that mirrors the release from competitive pressures. For example, the absence of larger carnivores in the private land can drive carnivore co-occurrence and activity patterns as opposed to the management strategies of the reserve. In the Brazilian Atlantic Forest, margay activity patterns remained consistent across different disturbance levels47. However, smaller mammals exhibited shifts in activity along a human-pressure gradient that reflected the absence of tigers (Panthera tigris) and leopards (Panthera pardus) in Malaysia48. In Peru, ocelots reduced diurnal activity in mixed-used areas compared to the PAs49. We found that grey foxes were more diurnal on private lands with activity peaking during the middle of the day but remained nocturnal in buffer zones where human detections were highest. Our findings were consistent with expectations for grey fox, as these ‘human adapters’ commonly use areas in or adjacent to human development50 compared to the more elusive margay that often prefer less disturbed ecosystems51. Previous works provide evidence that mesocarnivore behavior regarding interactions with apex predators can be meditated by the presence of co-occurring humans in shared landscapes[e.g., 52,53.
Coexistence is often facilitated by competing carnivores partitioning habitat that results in fine-scale avoidance behaviors and resource acquisition tactics in undisturbed landscapes[e.g., 54,55. Contrary to our expectations, activity overlap between the margay and ocelot decreased along the land-use gradient with the highest overlap in the core zones and lowest in private lands. The higher activity overlap in core zones may indicate that ocelots and margays easily coexist naturally since ocelots have larger home ranges and margay are able to seek refugia in trees given their increased arboreality51,56. Greater activity overlap in the buffer zones between competitor species may be driven by a combination of top-down pressures as both large carnivore and human detections were highest in the buffer compared to core and private land-use designations. Concerning resource acquisition, we found significant differences in activity levels between core zones and private lands with higher activity overlap in the buffer zones for all three predator-prey pairs (ocelot-armadillo, ocelot-opossum, margay-opossum). This finding contradicts expectations posited by the ‘human shield hypothesis’, where human presence and development act as a spatial refugia for prey that can limit their predation risks57,58. Instead, the increased temporal overlap in areas with high human activity corroborate mounting evidence of how anthropogenic disturbances reduce niche space for wildlife12,59,60.
Findings from our study elucidate concerns on how future PA designations may affect carnivore persistence in shared landscapes. However, additional sampling is needed to provide specific recommendations to aid in promoting coexistence, as our study was confined to a single season over five months, despite high sampling effort. We recognize broad temporal inferences should not be drawn from our work given the importance of seasonality in shaping carnivore diel activity and temporal overlap61. Additionally, our study relied on land-use designations as proxies for factors such as habitat quality, prey availability, and human activity. The surprising result of increased human activity in the buffer zones challenges assumptions about the role of land-use designations in shaping carnivore behavior. While buffer zones are often envisioned as transitional areas, in reality, they may serve as hotspots of human-carnivore interaction within REBITRI due to shared use. This unexpected pattern suggests that buffer zones, while intended to minimize human-wildlife conflict, might carry a higher burden of human disturbance compared to core zones or private lands, where human presence was relatively lower. It is also important to note that private lands in REBITRI are typically well-guarded and patrolled by landowners, leading to less human presence despite being outside PA boundaries. Private lands in our study were also treated as a homogeneous category, despite their inherent heterogeneity, ranging from large-scale commercial agriculture to small-scale rural holdings. More targeted studies examining direct measures of habitat quality and human influence would be needed to fully understand the dynamics driving these patterns. Given the frequent conflict between ecosystem protection, biodiversity conservation, and human livelihoods in PAs of the Sierra Madre de Chiapas region, including in REBITRI62, future studies should also explicitly include social indicators to comprehensively depict the complex socio-ecological system of REBITRI.
Our study underscores the potential of multiple-use landscapes to foster carnivore-human coexistence yet highlights the necessity for adaptive management strategies amidst shifting carnivore activity patterns under anthropogenic pressures. Effective management, as seen in mixed-use landscapes, involves community engagement and balanced conservation efforts7,63. As PAs adopt inclusive conservation models like OECMs in Mexico and beyond, prioritizing multiple-use landscape management will be vital for biodiversity conservation while addressing human needs64. Our findings in REBITRI emphasize the role of such landscapes in mitigating human impacts on carnivore behavior and biodiversity. However, managing human-wildlife conflicts and ensuring conservation sustainability pose ongoing challenges. By refining our understanding of activity dynamics, we can better inform conservation efforts aimed at ensuring the persistence of carnivores in increasingly human-dominated landscapes.
Data availability
Data generated or analysed during this study are included in this published article (and its Supplementary Information files). However, some data are sensitive corresponding to geographic ___location of sensitive species and mammal occurrences acquired from private lands that are restricted. Contact [email protected] as corresponding author for additional information to access data.
References
Ripple, W. J. et al. Status and ecological effects of the World’s largest carnivores. Science 343 (6167), 1241484 (2014).
Rabinowitz, A. & Zeller, K. A. A range-wide model of landscape connectivity and conservation for the jaguar, Panthera onca. Biol. Conserv. 143, 939–945 (2010).
Gray, C. L. et al. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 7 (1), Article1 (2016).
Ceballos, G. Conservation priorities for mammals in megadiverse Mexico: The efficiency of reserve networks. Ecol. Appl. 17 (2), 569–578 (2007).
Jackson, P. & Nowell, K. Wild cats: Status survey and conservation action plan. In IUCN (1996).
Bauer, H. et al. Lion (Panthera leopopulations are declining rapidly across Africa, except in intensively managed areas. Proc. Natl. Acad. Sci. 112 (48), 14894–14899 (2015).
Cumming, G. S. et al. Understanding protected area resilience: A multi-scale, social-ecological approach. Ecol. Appl. 25 (2), 299–319 (2015).
Palomo, I. et al. Incorporating the social–ecological approach in protected areas in the anthropocene. BioScience 64 (3), 181–191 (2014).
Geldmann, J. et al. A global-level assessment of the effectiveness of protected areas at resisting anthropogenic pressures. Proc. Natl. Acad. Sci. 116 (46), 23209–23215 (2019).
Jones, K. R. et al. One-third of global protected land is under intense human pressure. Science 360 (6390), 788–791 (2018).
Doherty, T. S., Hays, G. C. & Driscoll, D. A. Human disturbance causes widespread disruption of animal movement. Nat. Ecol. Evol. 5 (4), 513–519 (2021).
Cox, D. T. C., Gardner, A. S. & Gaston, K. J. Diel niche variation mammalian declines anthropocene. Sci. Rep. 13(1): 1031. (2023).
Harris, N. C. et al. First camera survey in Burkina Faso and Niger reveals human pressures on mammal communities within the largest protected area complex in West Africa. Conserv. Lett., 12(5) (2019).
Fehlmann, G. et al. Behavioral causes, ecological consequences, and management challenges associated with wildlife foraging in human-modified landscapes. BioScience 71 (1), 40–54 (2020).
Tucker, M. A. et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 359 (6374), 466–469 (2018).
Suraci, J. P. et al. Fear of humans as apex predators has landscape-scale impacts from mountain lions to mice. Ecol. Lett. 22 (10), 1578–1586 (2019).
Di Minin, E. et al. Global priorities for national carnivore conservation under land use change. Sci. Rep. 6 (1), 23814 (2016).
Hoesen, J. et al. Assessing the effectiveness of potential protected areas and OECMs in conserving biodiversity against subsurface resource extraction impacts. Biol. Conserv. 283, 110134 (2023).
Bezaury-Creel, J. & Gutiérrez-Carbonell, D. Áreas naturales protegidas y desarrollo social en México (Vol. 2, pp. 385–431.). Capital natural de Mexico, 2, 385–431 (2009).
CONANP, S. R. México se compromete a impulsar la conservación del 30% del planeta para el gob.mx (2030). Retrieved from http://www.gob.mx/sre/prensa/mexico-se-compromete-a-impulsar-la-conservacion-del-30-del-planeta-para-el-2030?idiom=es. (2020).
Anaya-Zamora, V., López-González, C. A. & Pineda-López, R. F. Factors associated with human-carnivore conflict in a protected area in central Mexico. Ecosistemas Y Recursos Agropecuarios. 4, 381–393 (2017).
Luna-Vega, I. et al. Threatened trees characteristic of Mexican tropical montane cloud forests. Diversity 15 (1), Article1 (2023).
Moguel, P. & Toledo, V. M. Biodiversity conservation in traditional coffee systems of Mexico. Conserv. Biol. 13 (1), 11–21 (1999).
Sévêque, A. et al. Human disturbance has contrasting effects on niche partitioning within Carnivore communities. Biol. Rev. 95 (6), 1689–1705 (2020).
Patten, M. A., Burger, J. C. & Mitrovich, M. The intersection of human disturbance and diel activity, with potential consequences on trophic interactions. PLoS ONE. 14 (12), e0226418 (2019).
Burton, A. C. et al. Mammal Responses to Global Changes in Human Activity Vary by Trophic Group and Landscape (Nature Ecology & Evolution, 2024).
Gallo, T. et al. Mammals adjust diel activity across gradients of urbanization. eLife 11, e74756 (2022).
Mills, K. L. & Harris, N. C. Humans disrupt access to prey for large African carnivores. eLife 9, e60690 (2020).
Lee, S. X. T. et al. Effects of human disturbances on wildlife behaviour and consequences for predator–prey overlap in Southeast Asia. Nat. Commun. 15 (1), 1521 (2024).
Manlick, P. J. & Pauli, J. N. Human disturbance increases trophic niche overlap in terrestrial carnivore communities. Proc. Natl. Acad. Sci. 117(43), 26842–26848 (2020).
Gittleman, J. L. et al. Carnivore Conservation (Cambridge University Press, 2001).
Balbuena-Serrano, Á. et al. Connectivity of priority areas for the conservation of large carnivores in northern Mexico. J. Nat. Conserv. 65, 126116 (2022).
Miguel Angel Pérez, F. & Vovides, A. P. Spatial distribution, population structure, and fecundity of Ceratozamia Matudai Lundell (Zamiaceae) in El Triunfo Biosphere Reserve, Chiapas, Mexico. Bot. Rev. 70 (2), 299–311 (2004).
Martínez-Meléndez, N., Pérez-Farrera, M. & MartÍnez-Camilo, R. The vascular epiphyte flora of El Triunfo Biosphere Reserve, Chiapas, México. Rhodora 111 (948), 503–535 (2009).
van der Hoek, Y. et al. Diversity and diel activity patterns of terrestrial mammals in the Nkuba Conservation Area, Democratic Republic of the Congo. Oryx 57, 107–117 (2023).
Kolowski, J. M. & Forrester, T. D. Camera trap placement and the potential for bias due to trails and other features. PLoS ONE. 12 (10), e0186679 (2017).
Greenberg, S., Godin, T. & Whittington, J. Design patterns for wildlife-related camera trap image analysis. Ecol. Evol. 9 (24), 13706–13730 (2019).
Niedballa, J. et al. camtrapR: An R package for efficient camera trap data management. Methods Ecol. Evol. 7 (12), 1457–1462 (2016).
Buchholz, R., Stamn, J. & Neha, S. A. Can camera traps be used to measure climate change induced alterations of the activity patterns of elusive terrestrial vertebrates? Clim. Change Ecol. 2, 100020 (2021).
Clauss, M., Scriba, M., Kioko, J., Ganzhorn, J. U. & Kiffner, C. Camera-trap data do not indicate scaling of diel activity and cathemerality with body mass in an East African mammal assemblage. Ecol. Evol. 11(20), 13846–13861 (2021).
Ridout, M. S. & Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 14, 322–337 (2009).
Meredith, M. & Ridout, M. S. The Overlap Package (2014).
Cox, D. T. & Gaston, K. J. Ecosystem Functioning Across the diel Cycle in the Anthropocene (Trends in Ecology & Evolution, 2023).
Vallejo-Vargas, A. F. et al. Consistent diel activity patterns of forest mammals among tropical regions. Nat. Commun. 13 (1), 7102 (2022).
Terborgh, J. et al. The Role of Top Carnivores in Regulating Terrestrial Ecosystems. Continental conservation: Scientific foundations of regional reserve networks, 39–64 (1999).
Ritchie, E. G. & Johnson, C. N. Predator interactions, mesopredator release and biodiversity conservation. Ecol. Lett. 12 (9), 982–998 (2009).
Horn, P. E. et al. Margay (Leopardus wiedii) in the southernmost Atlantic Forest: Density and activity patterns under different levels of anthropogenic disturbance. PLoS ONE. 15 (5), e0232013 (2020).
Kobayashi, S. et al. Impacts of human activity on wild mammal detection rates and diel activity patterns in Endau-Rompin National Park, Malaysia. J. Trop. For. Sci. 36 (1), 80–90 (2024).
Zwicker, S. & Gardner, B. Moderate anthropogenic impacts alter temporal niche without affecting spatial distribution of ocelots in the Amazon rainforest. Biotropica 56 (4), e13346 (2024).
Rodriguez, J. T., Lesmeister, D. B. & Levi, T. Mesocarnivore landscape use along a gradient of urban, rural, and forest cover. PeerJ 9, e11083 (2021).
De Oliveira, T. G. Leopardus wiedii. Mamm. Species. 579, 1–6 (1998).
Gámez, S. & Harris, N. C. Living in the concrete jungle: Carnivore spatial ecology in urban parks. Ecol. Appl. 31 (6), e02393 (2021).
Haswell, P. M. et al. Fear, foraging and olfaction: How mesopredators avoid costly interactions with apex predators. Oecologia 187 (3), 573–583 (2018).
Vanak, A. T. et al. Moving to stay in place: Behavioral mechanisms for coexistence of African large carnivores. Ecology 94 (11), 2619–2631 (2013).
Miller, J. R. B. et al. Lions and leopards coexist without spatial, temporal or demographic effects of interspecific competition. J. Anim. Ecol. 87 (6), 1709–1726 (2018).
Gámez, S. & Harris, N. C. Conceptualizing the 3-dimensional niche and vertical space use. Trends Ecol. Evol. 37, 953–962 (2022).
Berger, J. Fear, human shields and the redistribution of prey and predators in protected areas. Biol. Lett. 3 (6), 620–623 (2007).
Gaynor, K. M., McInturff, A. & Brashares, J. S. Contrasting patterns of risk from human and non-human predators shape temporal activity of prey. J. Anim. Ecol. 91, 46–60 (2021).
Murphy, A. et al. Threading the needle: How humans influence predator–prey spatiotemporal interactions in a multiple-predator system. J. Anim. Ecol. 90 (10), 2377–2390 (2021).
Bonnot, N. C. et al. Fear of the dark? Contrasting impacts of humans versus lynx on diel activity of roe deer across Europe. J. Anim. Ecol. 89 (1), 132–145 (2020).
Smith, K. et al. Temporal partitioning and the potential for avoidance behaviour within South African Carnivore communities. Ecol. Evol. 13 (8), e10380 (2023).
Cortina-Villar, S. et al. Resolving the conflict between ecosystem protection and land use in protected areas of the Sierra Madre De Chiapas, Mexico. Environ. Manag. 49 (3), 649–662 (2012).
Boron, V. et al. The role of unprotected and privately protected areas for ocelot conservation: Densities in Colombia and Brazil. J. Mammal. 103 (3), 639–647 (2021).
Peña-Azcona, I. et al. Áreas de conservación voluntaria en México: Alcances y desafíos. Revista De Ciencias Ambientales. 56 (2), 120–145 (2022).
Acknowledgements
We would like to express our sincere gratitude to our local field technicians (N. Guzman, D. Najera, O. Domingues, and V. Valentin) and the four local communities we worked with (Tres de Mayo, Finca Custepec, Plan de Ayala, & Caballo Blanco) as they were essential to the project’s success. We’d like to thank property owners D. Marin, D. Domingo, D. Moises, and M. Guzman for letting us place camera traps on their property. We would also like to thank our government partners, the National Commission of Protected Natural Areas (CONANP) officials based in Mapastepec, and our university partners Dra. C. Lorenzo-Monterrubio and Biol. J. Bolaños Citalán at El Colegio de la Frontera Sur (ECOSUR) in San Cristobal de las Casas for their support. We thank the Tropical Resources Institute and the Council on Latin American and Iberian Studies at Yale University for their financial support. Gámez gratefully acknowledges financial support for this research by the Fulbright U.S. Scholar Program, which is sponsored by the U.S. Department of State and the U.S.-Mexico Commission for Educational Exchange (COMEXUS) and the Ford Foundation Fellowship. Finally, we acknowledge the invaluable support from lab members including S. Alinaeem and H. Pliske in the Applied Wildlife Ecology Lab at the Yale School of the Environment with project development and camera trap image sorting.
Author information
Authors and Affiliations
Contributions
GG and NCH conceived the study with all authors participating in the study design. GG and SG led field logistics and completed the data collection. GG processed and analyzed data as well as wrote a first draft of the manuscript. NCH supervised and provided financial support. All authors edited and gave final approval for the publication.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
González, G., Gámez, S. & Harris, N.C. Carnivore activity across landuse gradients in a Mexican biosphere reserve. Sci Rep 15, 4431 (2025). https://doi.org/10.1038/s41598-025-87850-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-87850-7
