Abstract
Through isotopic investigations of directly dated human remains recovered from the Eton College Rowing Course, we examine changes in diet from the Neolithic to the Roman period. The human isotope signatures point to a diet based on C3 terrestrial resources. A significant correlation is visible between human δ13C values and time, but no such trend is observed in δ15N. The animal isotope data from Eton are unevenly distributed, making it difficult to determine if the human values mirror the animal values. To assess whether the results from Eton are typical, we compare our results to isotope data from other British sites dating from the Neolithic to the Roman period. Across this time period, we see a strong correlation between the mean δ15N of the humans and that of the main domesticated herbivores, with an offset of ∼4.5‰ between the two. Thus, the changes in the human isotope values are likely linked to changes in the isotopic signatures of the herbivores rather than changes in the protein composition of human diets. By contrast, no clear temporal relationship is observed between the mean δ13C of the humans and that of the main domesticated herbivores, with an offset of ∼1.4‰ between the two. There is, however, a weak correlation observed between the mean δ13C of the humans and that of the cattle, which may account for some of the variation in human δ13C values between sites. The absence of a strong correlation between mean human and animal δ13C suggests that the primary factor influencing human δ13C values between sites is dietary composition. The lack of co-variation between δ13C and δ15N is likely to reflect the different representation of dietary macronutrients. Given that the nitrogen results suggest that the animal protein consumption patterns are similar across sites, the human δ13C variation between sites is likely to reflect the plant portion of the diet.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Geoarchaeological investigations of an area of the Thames flood plain were undertaken by Oxford Archaeology during the construction of the Eton College Rowing Course at Dorney, South Buckinghamshire (NGR SU 920770; Fig. 1). Situated on the north bank of the present channel of the Thames, the area of the flood plain investigated covered approximately 150 ha (Allen and Welsh 1996). The investigation focussed on a large palaeochannel of the Thames (which covered approximately half the site at one time or another), along with the adjacent alluvial floodplain and gravel terraces (Fig. 2). These contain archaeological material dating from the Upper Palaeolithic to the Mediaeval period (Allen et al. 2000; Parker et al. 2008). Human remains were recovered from the palaeochannel and from the gravel terraces, a selection of which have been radiocarbon dated to the Neolithic (ca. 4000–2500 bc), Bronze Age (ca. 2500–750 bc) and Iron Age (ca. 750 bc–ad 50).

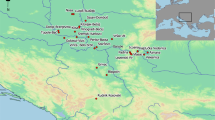
Location of Eton College Rowing Course and other sites mentions within paper. 1 Ascott Under Wychwood, 2 Hazleton, 3 Hambledon Hill, 4 Brean Down, 5 Cladh Hallan, 6 Irthingbourgh, 7 Gayhurst, 8 Hampshire (Micheldiver wood, Winnall Down), 9 Danebury, 10 Poundbury, 11 Glastonbury, 12–14 Cornwall (Trevelgue Head, Harlyn Bay, Trethellan Farm), 15 Wetwang, 16–18 East Lothian (Port Seton, Broxmouth, Dryburn Bridge), 19 York, 20 Gloucester, 21 Yarnton, 22 Queensford Farm, 23 Dorset, 24 Lismore

Map of Rowing Lake site showing the areas of archaeological exaction and watching brief, and labelling the main discoveries, including ring ditches (reproduced with the permission of Tarmac Ltd)
The environmental studies at Eton and at other areas of the Upper and Middle Thames Valley have provided both broad dietary information and landscape reconstructions (Lambrick 1992; Parker et al. 2008). These studies, however, provide only indirect qualitative information about the diet of the population occupying the area. Stable carbon and nitrogen isotope investigations of human remains can be used to obtain direct, quantitative, long-term records of past diets. Many of the human remains were recovered from palaeochannel deposits; thus, archaeological information directly linked to these individuals is limited. The first aim of this study has been to further illuminate understanding of the human remains recovered from the Eton College Rowing Course by reconstructing their diets through carbon and nitrogen isotopic analysis of bone collagen. The second aim has been to establish whether the diets of the Neolithic, Bronze Age and Iron Age people excavated from Eton were isotopically similar or different. The third aim is to establish whether there were isotopic differences in diet between the humans buried on land and those deposited in the river.
Reconstructing diet by stable isotope analysis
The carbon and nitrogen isotope signatures in bone collagen are thought to primarily reflect the protein consumed by an individual (Ambrose and Norr 1993). Isotopic analysis of bone collagen can provide information about the long-term diet of an individual as bone turnover is relatively slow; thus, its collagen isotopic signatures reflect the dietary protein consumed over a period of years (Hedges et al. 2007; Stenhouse and Baxter 1979). Carbon isotope signatures are often used to detect the type of plants consumed (C3 versus C4 plants) by an individual (e.g. Vogel and Van der Merwe 1977). During the time periods investigated in this study, however, C4 plants are unlikely to have made any significant contribution to diet of humans in the British Isles. Carbon isotope signatures can also be used to detect variations in the amount of marine versus terrestrial protein consumed (e.g. Lightfoot et al. 2011; Schoeninger et al. 1983). An individual’s nitrogen isotope signature reveals their position in the food chain because an enrichment of around +3‰ to +5‰ occurs with each increase in trophic level (Bocherens and Drucker 2003). This amount of trophic level enrichment has been observed between human and animal bone collagen values from Neolithic to post-Roman sites in Northwest Europe (Hedges and Reynard 2007). Therefore, herbivores, omnivores and carnivores can be distinguished via nitrogen isotope analysis. As marine and freshwater food chains are longer than terrestrial ones, consumption of meat can be distinguished from consumption of marine and freshwater fish (Katzenberg and Weber 1999; Richards and Hedges 1999). Minor component of an individual’s diet may not be detected using isotopic techniques as more major components may mask their isotopic input. For example, small amounts of freshwater proteins could be masked by the consumption of young animals (Boric et al. 2004). Climatic and environmental parameters can influence plant carbon and nitrogen isotope signatures, which are then passed up the food chain (Stevens and Hedges 2004; Stevens et al. 2008). Thus, it is essential to sample contemporaneous fauna along with the humans at a site in order to determine the isotopic signatures of food sources potentially consumed by the human population.
Archaeobotanical and archaeozoological evidence for human subsistence in the Middle/Upper Thames Valley
The evidence for human subsistence in the Middle and Upper Thames Valley from the Neolithic to the Roman period has been recovered during the excavation of a number of farmsteads. A summary of the plant and animal remains found during excavation of Middle/Upper Thames valley sites is given in Table 1. During the Neolithic, pastoralism was paramount and residue analysis shows that dairying played a role in the local economy from the beginnings of agriculture (Copley et al. 2003). Animal bone assemblages show the importance domesticated compared to wild species (Serjeantson 1996), while plant assemblages are dominated by wild plant foods (Moffett et al. 1989). Mixed farms and specialisation developed by the Late Bronze Age (Bradley et al. 1980; Campbell 1992; Lambrick 1992; Lewis and Batt 2006). At the end of the Late Bronze Age, there was a significant settlement shift in the Middle Thames Valley, and fewer early Iron Age settlements have been excavated. Nevertheless, the range and relative importance of the various animal species was similar. The traditional crops, emmer wheat and naked barley, went into marked declined, the latter disappearing entirely, and were replaced by spelt wheat and hulled barley (Jones 1984). Only limited use was made of wild food crops, such as hazelnut and berries (Booth et al. 2007). During the Roman period, there was a high population density in the Middle Thames Valley, with intensification of farming (as occurred at Eton College Rowing Course) and the introduction of many new crop species. The major cereal crops and domesticated animals, however, show continuation from the Iron Age. As the major domesticated animal and plants do not change substantially between the periods studied, we expect any isotopic variation over time to be minor.
Human remains at Eton College Rowing Course
Carbon and nitrogen isotope analysis was carried out on skeletal remains from 19 humans recovered from the Eton College Rowing Course. All but one of these humans had been previously radiocarbon dated. Six of the humans were radiocarbon dated to the Neolithic, seven to the Bronze Age, five to the Iron Age and one to the Roman period. Thirteen of the humans sampled for isotope analysis were recovered from palaeochannels of the River Thames, and six were recovered from dry-land contexts. The Neolithic human bone assemblage at the site represents a wide range of disposal practices including formal inhumation, the deliberate placing of skulls (and possibly whole bodies) into the river and the incorporation of human bones into pits and middens (Allen et al. 2000).
The Neolithic inhumations consisted of a crouched burial of an adult female (about 25–35 years old) and an infant (about 5–6 years old), which were located just outside a ring ditch (Allen et al. 2000). Radiocarbon dates on human remains found in the palaeochannel indicate that a further four individuals date to the Neolithic (Fig. 2, Table 2). The Bronze Age human assemblage consisted of three crouched inhumations. The burials of a male adult (about 35 years old) and a juvenile (about 12 years old) were located on the edge of a Bronze Age watering hole. Deveral-Rimbury pottery was recovered from the fill of the former burial. The third Bronze Age crouched burial was cut into a ring ditch (which contained sherds of Beaker pottery) and belonged to a female (about 18–20 years old), who was found with a foetus and therefore is thought to have either been pregnant at death or to have died in child birth (Allen et al. 2000). All but one of the Bronze Age human remains recovered from palaeochannel contexts were located on a sandbank. It is possible that these remains derived from burials on dry-land upriver; however, there is evidence that the sandbanks were being visited and used for ritual purposes (Allen et al. 2000). None of the Iron Age humans sampled for isotopic analysis were from inhumation burials; all were recovered from palaeochannel deposits. Thus, their association with the Iron Age farmstead and settlement found within the excavation area is unclear. Bones of three of the five Iron Age humans were found in close proximity to three wooden structures interpreted as bridges (Allen et al. 2000). Finding more than one bone from a single individual, including one case of articulating bones, downstream of one bridge makes redeposition from graves on the bank further upstream unlikely (Allen et al. 2000). These bones could represent people who drowned or whose bones were placed in the river soon after death. The single Roman burial was an inhumation and was associated with the Roman farmstead. Allen et al. (2000) recorded the age and sex profile of the human remains if diagnostic skeletal elements were available. In this study, we use their determinations of age and sex for comparison with isotopic results.
Material and methodology
Samples were obtained from the bones of 19 humans dating to the Neolithic, Bronze Age, Iron Age and Roman period. In addition, bone samples were also taken from a range of contemporaneous fauna; 15 cattle, 14 sheep, 12 pigs, 11 horse, three red deer and one roe deer. Full details for specimens selected for sampling are listed in Table 2. Sampling focussed on adult specimens; young and juvenile animals were avoided. Samples were prepared following the method in Privat et al. (2002). With palaeochannel deposits covering around half of the site, it is very hard to firmly attribute fauna to specific archaeological periods; therefore, the number of samples analysed for each species from each time period is relatively small (Table 3). A further six isotopic results were obtained for animals during radiocarbon dating: four cattle, one horse and one red deer (Table 3). Collagen was extracted at the RLAHA, Oxford. Samples were isotopically analysed using an automated Carlo Erba carbon and nitrogen elemental analyser coupled in a continuous flow mode to an isotope ratio-monitoring (PDZ Europa Geo 20/20) mass spectrometer. Carbon and nitrogen results are measured in parts per mille (‰) relative to VPDB and AIR standards, respectively (Hoefs 1997). Each sample was run at least in duplicate. Replicate measurement errors on laboratory standards (comprising in-house standards of nylon and alanine calibrated against IAEA standards) were <0.2‰ over the period of analysis. For the six 14C dated animals, the analytical errors for the δ13C and δ15N results are larger, potentially as large as 0.3‰ (Peter Ditchfield, personal communication).
Results and discussion
Sample preservation
Thirteen out of the 81 samples failed to produce enough collagen for isotope analysis. The successfully extracted collagen had C/N atomic ratios between 2.9 and 3.6 (Tables 2 and 3). This is considered to be indicative of good collagen preservation (DeNiro 1985). The δ13C and δ15N values of the humans and fauna analysed are listed in Tables 2 and 3.
All periods
Human values range from −21.4‰ to −19.7‰ in δ13C with a mean of −20.5‰ (σ = 0.5‰) and from 8.9‰ to 13.0‰ in δ15N with a mean of 10.7‰ (σ = 1.0‰). The associated animals range from −22.8‰ to −19.3‰ in δ13C with a mean of −21.8 (σ = 0.7‰) and from 3.9‰ to 10.1‰ in δ15N with a mean of 6.3‰ (σ = 1.5‰). A summary of the human and animal isotope results can be found in Table 4. The human δ13C and δ15N signatures point to a diet based on C3 terrestrial resources. There is no evidence of consumption of C4 plants or marine resources among the Eton individuals analysed. The highest human δ15N values might indicate a very small contribution of freshwater fish to the diet; however, despite extensive sieving, fish bones were only recovered from the palaeochannel at Eton College Rowing Course. No fish bones (with the exception of a single pike bone within Neolithic grave 5888) were recovered outside of the palaeochannel. It is unlikely that this absence of fish bones in terrestrial contexts is a result of taphonomic factors. Therefore, we conclude that fish were at most a very minor dietary component.
The number of humans from each time period is limited, which constrains the strength of the statistical analyses. Nevertheless, some statistically significant observation can be made. The human δ15N values do not appear to change through time (Figs. 3 and 4) unlike the human δ13C values (Figs. 3 and 4). The mean δ13C of the Neolithic humans is significantly different to those of the Bronze Age and Iron Age humans (one-way ANOVA with post-hoc Bonferroni correction, p = 0.001 and p ≤ 0.001, respectively); however, those of the Bronze Age and Iron Age humans are not significantly different to each other (Fig. 3). With only one Roman individual, it is not possible to say whether or not the Roman human δ13C value is significantly different to those of humans from other time periods. When the Neolithic, Bronze Age and Iron Age human δ13C values are plotted against uncalibrated radiocarbon date bp, a highly significant correlation is visible (R 2 = 0.7169; Fig. 4). The human δ13C values from the Bronze Age cluster into two groups (Fig. 4). Interestingly, the two higher δ13C values (specimen A/ERL/B/1 and A/ERL/B/15) were found together, next to a Bronze Age watering hole. When the carbon and nitrogen isotope signatures of the Bronze Age and Iron Age humans are plotted together, they do seem to be distinct however the differences between the mean δ13C and δ15N values of the two groups are not statistically significant.

Eton College Rowing Course human and animal δ13C and δ15N bone collagen isotope signature plotted by period (Neo Neolithic, BA Bronze Age, IA Iron Age, Rom Roman)

δ13C and δ15N of Eton College Rowing Course humans over time. A positive correlation is seen between δ13C and uncalibrated radiocarbon date bp for Neolithic, Bronze Age and Iron Age humans
To determine whether variation in human carbon isotope signature over time is due to changing dietary intake or variations in the isotopic signature of the same diet, it is necessary to investigate whether the isotope signatures of the animals consumed by the humans varied over time. The number of associated animals available for isotopic analysis was limited due to problems of securely linking animal remains to a specific time period, as many of the animals were recovered from palaeochannel contexts and thus may have been redeposited. Due to the limited number of sheep and red deer, it is not possible to say whether there are any temporal changes in their isotope signatures. The isotope signatures of the horses did not change extensively between the Bronze Age and Roman period (Fig. 3).
More variation is seen in the cattle isotope signatures over time. The mean cattle δ15N is lower in the Neolithic than in the Bronze Age (Table 4), and this difference is significant (p = 0.004). The Bronze Age cattle δ15N could be elevated due to the consumption of plants that had been grown on soils that have been fertilised with manure. This could indicate a change in cattle herd management practices. The mean cattle δ15N in the Iron Age and Roman period are lower than in the Neolithic and Bronze Age; however, the difference is only statistically significant between the Bronze Age and the Iron Age (p = 0.002). This variation in cattle δ15N values is not mirrored in the human δ15N values. There is a small amount of variation in the mean cattle δ13C over time, which is not significant but broadly follows this same trend as the human δ13C values.
With only a single pig data point from the Bronze Age and none from the Iron Age, little can be said about the nature of their diet during these time periods. A substantial shift in both pig δ13C and δ15N values can be seen, however, between the Neolithic and Roman period. Mean pig δ13C and δ15N are significantly lower and higher respectively during the Roman period than during the Neolithic (Table 4, p = 0.002 and p = 0.018, respectively). This trend in δ13C values is the opposite to that observed in human δ13C values. The positive pig δ13C values during the Neolithic has been noted elsewhere and has been linked to the use of wildwood resources, particularly fungi, during this period (Hamilton et al. 2009). The elevated δ15N values found in the Roman pigs is indicative of an omnivorous diet most likely including meat scraps or excreta from household waste.
The quantity of the baseline animal isotopic data from Eton against which the human δ13C and δ15N values can be compared is limited and unevenly distributed across species and time periods. It is therefore hard to establish for certain whether the change in human δ13C values relates to change in types of food consumed or change in the isotope signatures of the foods consumed. Through comparison of the Eton isotope data with that from other British sites, it may be possible to further illuminate our understanding of the trend observed in human and animal isotope signatures at Eton.
Neolithic
The Neolithic human and animal isotope signatures from the following sites have been published: YarntonFootnote 1 (Lightfoot et al. 2009) and Ascott Under Wychwood (Hedges et al. 2006) (located in the Thames Valley) and Hazelton (Gloucester) (Hedges et al. 2008) and Hambledon Hill (Dorset) (Richards 2008) (non-Thames Valley sites). The isotope values of the humans from these sites are plotted in Fig. 5 along with the animal isotope signatures from the Thames Valley sites only. Data from non-Thames Valley Neolithic animals are not included in Fig. 5 (as their inclusion makes the figure too cluttered) but are however listed in electronic supplementary data file.

Human and animal δ13C and δ15N bone collagen isotope signature from Neolithic sites: Eton College Rowing Course, Ascott Under Wychwood (Hedges et al. 2006), Hambledon (Richards 2008), Hazelton (Hedges et al. 2008) and Yarnton (Lightfoot et al. 2009). Only animals from Eton, Yarnton and Ascott Under Wychwood are including in graph. See Electronic supplementary data file for animal isotope results from other Neolithic sites
The δ13C of the Neolithic humans at Eton are similar to those of human from the Neolithic sites mentioned above. Their mean δ15N is somewhat higher than that of the humans from Ascott Under Wychwood, Hazleton and Hambledon Hill; however, this difference is statistically significant (one-way ANOVA with post hoc Bonferroni correction: p = 0.024, p = 0.002 and p ≤ 0.001, respectively). Although the mean δ15N of the Eton humans is lower than that of the Yarnton humans they are not significantly different (p = 0.127). It should be noted however that there are only a small number of Neolithic human samples from Eton and Yarton. Like at Eton, the mean δ15N of the Yarnton humans is significantly higher than that of the humans from Ascott Under Wychwood, Hazelton and Hambledon Hill (p = 0.002, p ≤ 0.001 and p ≤ 0.001, respectively; see footnote 1). From the data, it appears that Neolithic humans from Yarnton and Eton have elevated δ15N values relative to contemporaneous humans from other areas of the British Isles. This pattern is partially repeated in the pig δ15N values, which are higher at Eton than at Hazelton and Hambledon Hill, but are similar to those at Ascott Under Wychwood. The pattern is not seen in the cattle whose isotope signatures are similar in both Thames Valley and non-Thames Valley sites. The single Neolithic sheep from Eton has isotope values that are broadly similar to those of sheep at Ascott Under Wychwood, but no real interpretation can be made due to the limited data. Mean pig δ13C at Eton is more positive than other herbivores, suggesting their greater use of wildwood resources, including fungi (see Hamilton et al. 2009).
Bronze Age
Previous studies investigating the isotope signatures for Bronze Age humans and animals in the British Isles are very limited. “The Beaker Isotope Project” currently being lead by the University of Sheffield will greatly change this situation. Previous studies of Bronze Age human isotope signatures include that from Yarnton (see footnote 1) (Lightfoot et al. 2009) (Thames Valley site), and Cladh Hallan (Parker Pearson et al. 2005), Brean Down (Britton et al. 2008), Gayhurst and Irthingborough (Towers et al. 2011) and other sites in Scotland and Yorkshire (Jay and Richards 2007a) (Fig. 6, Electronic supplementary data file).

Human and animal δ13C and δ15N bone collagen isotope signature from Bronze Age sites: Eton College Rowing Course, Yarnton (Lightfoot et al. 2009), Cladh Hallan (Parker Pearson et al. 2005), Yorkshire (Jay and Richards 2007a), Scotland (Jay and Richards 2007a), and Brean Down (Britton et al. 2008). See Electronic supplementary data file for results
The mean Eton human δ15N value is similar to that of the Yarnton humans, whereas the mean δ13C is slightly more positive; however, this difference is not statistically significant (p = 0.051). Both the δ13C and δ15N values of the Eton humans are more positive than those from Yorkshire and Scotland (Jay and Richards 2007a). As associated Bronze Age faunal isotope data are limited from Eton and absent from the sites studied by Jay and Richards (2007a) and Lightfoot et al. (2009), it is not possible to say whether variation in human isotope values between sites relate to difference in the types of foods consumed or difference in the baseline isotopic signatures. Eton human mean δ15N is slightly higher than that of the humans from Cladh Hallan, whereas their mean δ13C values are similar. The larger standard deviation at Cladh Hallan suggests that the Cladh Hallan population had a more varied diet than the Eton population. The comparability of the two sites is however questionable as, although both in the British Isles, they are around 500 miles apart and the burial context differ tremendously as the Cladh Hallan remains are mummified (Parker Pearson et al. 2005). A small amount of Bronze Age animal isotope data are also available from Brean Down (Britton et al. 2008), where elevated sheep and cattle δ15N values were attributed to grazing on salt marshes and from Gayhurst and Irthingborough (Towers et al. 2011). Interestingly, the Brean Down cattle δ15N values are similar to those of Bronze Age cattle from Eton (Fig. 6). The Gayhurst and Irthingborough cattle have much lower δ13C and δ15N than those from Brean Down and Eton, but are more similar to the Neolithic, Iron Age and Roman cattle at Eton (see supplementary dataset). The Eton cattle δ15N could be elevated due to them consuming plants that have been grown on soils that have been fertilised with manure.
Iron Age
In comparison to the preceding periods, isotopic techniques have been applied to a reasonable number of Iron Age archaeological sites including Yarnton (Lightfoot et al. 2009) (Thames Valley site), Hampshire (Jay and Richards 2007b), Glastonbury (Jay 2008), Wetwang (Jay and Richards 2006), Cornwall (Jay and Richards 2007b), East Lothian (Jay and Richards 2007b), Poundbury (Richards et al. 1998) Dorset (Redfern et al. 2010), Danebury (Lightfoot and Stevens 2012; Stevens et al. 2010) and Lismore (animals only; O’Connell and Kimball 2006). Humans from all sites are included in Fig. 7 along with animals from Thames Valley sites (Eton and Yarnton). Animal isotope results from other Iron Age sites can be seen in the Electronic supplementary data file.

Human and animal δ13C and δ15N bone collagen isotope signature from Iron Age sites: Eton College Rowing Course, Yarnton (Lightfoot et al. 2009), Hampshire (Jay and Richards 2007b), Glastonbury (Jay 2008), Wetwang (Jay and Richards 2006), Cornwall (Jay and Richards 2007b), East Lothian (Jay and Richards 2007b), Poundbury (Richards et al. 1998), Dorset (Redfern et al. 2010) and Danebury (Lightfoot and Stevens 2012; Stevens et al. 2010). Only animals from Eton and Yarnton are including in graph. See Electronic supplementary data file for animal isotope results from other Iron Age sites
The Eton human δ13C values are similar to those from other Iron Age sites. Eton human isotope signatures are most similar to those from Yarnton and Dorset; however, limited variation is visible within the Iron Age human isotope signatures, with <1‰ variation occurring between the mean human δ13C values. The human δ15N values are similar to those from all of the above mentioned Iron Age sites, with the exception of those from Hampshire, Poundbury and Danebury, which are lower in δ15N. This difference appears to be due to the lower baseline values (in plants and passed on to the animals) due to local environmental conditions. This highlights the need to compare human isotope values to those of local fauna wherever possible. The isotope results are consistent with an Iron Age diet that was high in animal protein. However, recent research has shown that at some Iron Age sites cereal grain δ15N values can be similar to those of herbivores, making it difficult to determine the amount of animal protein in the diet (Lightfoot and Stevens 2012).
Roman period
Only one human from Eton College Rowing Course dating to the Roman period was sampled for isotope analysis, although associated animals were more abundant and can be compared to those from other Roman sites in the British Isles (Fig. 8, electronic supplementary data file); Yarnton (Lightfoot et al. 2009), Queenford Farm (Fuller et al. 2006) (both Thames Valley sites), and Poundbury (Richards et al. 1998), Gloucester (Chenery et al. 2010), Dorset (Redfern et al. 2010) and York (Müldner and Richards 2007). The δ13C and δ15N values of the Roman human from Eton are generally similar to those of humans analysed from the Roman sites mentioned above. The exception to this is the “high status” humans at Poundbury, whose δ13C values are higher than those at other Roman sites, most likely due to the consumption of fish. Some variation is visible in the human δ15N values, but these do not appear to relate to differences in dietary intake, rather due to differences in the baseline isotope values. For example, at Yarnton the human δ15N values are elevated; however, the associated animals also have elevated δ15N values.

Human and animal δ13C and δ15N bone collagen isotope signature from Roman sites: Eton College Rowing Course, Yarnton (Lightfoot et al. 2009), Queenford Farm (Fuller et al. 2006), Poundbury (Richards et al. 1998), Gloucester (Chenery et al. 2010) and York (Müldner and Richards 2007). Only animals from Eton and Yarnton are including in graph. See electronic supplementary data file for animal isotope results from other Roman sites
Relationship between human and faunal δ13C and δ15N variation
The comparisons made above have suggested that the human δ13C and δ15N values broadly track changes in the isotope values of the local fauna rather than changes in dietary practice. In order to further elucidate this relationship, the mean human isotope values at each site were compared to that of the main domesticated herbivores (cattle, sheep and horse) at each site.
A good correlation (R 2 = 0.6833) is seen between the mean δ15N of the humans and that of the main domesticated herbivores (Fig. 9). This confirms that animal δ15N values are the main control on the δ15N of the humans. Changes in the human isotope values during these periods are linked to changes in the isotopic signatures of the herbivores rather than changes in protein composition of human diets (e.g. consumption of freshwater resources). Two possible scenarios could result in changes in the herbivore diet isotope signatures. The variation in local environmental conditions could influence the isotopic values of the plants consumed by the herbivores, thus resulting in spatial and temporal variations being passed up the foodchain. Alternatively, changes in herbivore δ15N could be linked to changes in isotopic values and/or composition of animal diet due to varying farming methods such as manuring, irrigating or changing husbandry practices. Notably, the offset between mean human and mean herbivores δ15N values is 4.5‰ (σ = 0.6‰) with no sizeable changes in this offset between archaeological time period. This is consistent with that observed by other authors (e.g. Bocherens and Drucker 2003).

Mean human δ15N values from Neolithic to Roman sites in the British Isles plotted versus mean main domesticate (cattle, horse, sheep) δ15N values from the same sites
There is no clear relationship between the mean δ13C of the humans and that of the main domesticated herbivores (R 2 = 0.1327). The human δ15N values track the animal δ15N values closely because collagen nitrogen only comes from the protein component of the diet (Schoeller 1999). By contrast, the collagen carbon reflects primarily the dietary protein, although there is a contribution from carbohydrates and fats (Ambrose and Norr 1993; Fogel and Tuross 2003; Jim et al. 2006; Tieszen and Fagre 1993), which may explain the lack of co-variance between the humans and the animals. The highest correlation is observed between the mean δ13C of the humans and that of the cattle (Fig. 10), although the relationship is weak (R 2 = 0.3643). Thus, changes in the human δ13C values between sites may be partly due to changes in the isotopic composition of the cattle they consumed. As for the nitrogen, the weak correlation between cattle and human δ13C could reflect natural changes or anthropogenic changes in the isotopic composition of the cattle diet. The absence of a strong correlation between mean human and animal δ13C suggests that the primary factor influencing the differences in human δ13C values between sites is human dietary composition. Given that the nitrogen isotope values suggest that the animal protein consumption patterns are similar across sites, the human δ13C variation between sites is likely to reflect the plant portion of the diet. This could be due changes in the plant δ13C values, changes in the plant types consumed or changes in the proportion of plants and animals in the diet. The first two are unlikely to be the primary control on the variation as either of these scenarios would result in variation in the animal carbon isotope values (and therefore result in co-variation). Since the δ15N does vary between sites due to local ecological factors, it is likely that the carbon also does, although may not be fully detectable. Notably, the offset between mean human and mean herbivores δ13C values is 1.4‰ (σ = 0.5‰) with no sizeable changes in this offset between archaeological time period. This is consistent with that observed by other authors (e.g. Bocherens and Drucker 2003).

Mean human δ13C values from Neolithic to Roman sites in the British Isles plotted versus mean cattle δ13C values from the same sites
Conclusions
The isotopic analyses of human remain from the Eton College Rowing Course has confirmed that from the Neolithic to the early Roman period, the humans consumed a terrestrial diet from a C3 ecosystem. The contribution of marine or freshwater fish to the human diet was negligible. No significant differences were observed between the isotopic results of the humans buried on land and those deposited in the river, indicating that although their treatment in death differed, during life, their diets were likely to have been similar. No temporal trends in the human nitrogen isotope values were found, suggesting that diets during the Neolithic to the early Roman period at Eton did not alter substantially. Differences in mean human δ15N values were seen between sites in the British Isles; however, the high covariance between the mean human and main domesticate δ15N values indicate these differences relate to variation in local ecological conditions rather than dietary protein composition or amount of dietary protein consumed.
By contrast, a strong temporal trend was seen in the Eton human δ13C results, with values becoming more positive through time. Although animal samples are limited in number, the human δ13C values do not appear to directly mirror changes in the animal δ13C values, suggesting that the change in human δ13C over time at Eton may relate to a change in dietary composition. Differences in mean human δ13C values were seen between sites in the British Isles, but there is no clear relationship between the mean δ13C of the humans and that of the main domesticated herbivores, with only a weak covariance seen with cattle δ13C. Given that any changes in plant δ13C isotope values or plant type are likely to also be seen in animals, the absence of a strong correlation between mean human and animal δ13C suggests that changes in plant δ13C or plant type are not the primary cause of differences in human δ13C values between sites. While it is likely that there is variation in plant and animal δ13C between sites (due to either local environmental conditions or farming practices), we suggest that since there is no clear relationship between faunal collagen and human results the primary factor influencing the differences in human δ13C values between sites is dietary composition.
Notes
Note that Yarnton humans are Neolithic/Bronze Age.
References
Allen TG, Welsh K (1996) Eton College Rowing Course. Curr Archaeol 148:124–127
Allen TG, Hacking P, Boyle A (2000) Eton College Rowing Course: the burial traditions. Tarmac Papers 4:65–106
Allen T, Hayden C, Lamdin-Whymark H (2009) From Bronze Age enclosure to Anglo-Saxon settlement: archaeological excavations at Taplow hillfort, Buckinghamshire. Oxford Archaeology Thames Valley Landscapes Monograph No 30. Oxford Archaeological Unit, Oxford
Allen TG, Anderson E, Barclay A, Cromarty A-M, Knowles G, Lamdin-Whymark H, Parker AG, Robinson MA (2012a) Opening the wood, making the land. The archaeology of the Eton College Rowing Course and the Maidenhead, Windsor and Eton flood alleviation scheme, vol 1: Mesolithic to Early Bronze Age, Oxford Archaeology Thames Valley Landscapes Monograph. Oxford Archaeological Unit, Oxford (in press)
Allen TG, Bradley P, Parker AG, Robinson MA (2012b) Bridging the river, dividing the land: the archaeology of the Eton College Rowing Course and the Maidenhead, Windsor and Eton flood alleviation scheme, vol 2: Middle Bronze Age to Roman, Oxford Archaeology Thames Valley Landscapes Monograph. Oxford Archaeological Unit, Oxford (in press)
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe G (eds) Prehistoric human bone—archaeology at the molecular level. Springer, Berlin, pp 1–37
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Booth P, Dodd A, Robinson M, Smith A (2007) The Thames through time: the archaeology of the gravel terraces of the Upper and Middle Thames: the early historical period: AD1–1000. Oxford University School of Archaeology for Oxford Archaeology, Oxford
Boric D, Grupe G, Peters J, Mikic Z (2004) Is the Mesolithic–Neolithic subsistence dichotomy real? New stable isotope evidence from the Danube Gorges. Eur J Archaeol 7:221–248
Bradley RJ, Lobb S, Richards J, Robinson M (1980) Two late Bronze Age settlements on the Kennet gravels: Excavations at Aldermaston Wharf and Knights Farm, Burghfield, Burkshire. P Prehis Soc 46:217–295
Britton K, Müldner G, Bell M (2008) Stable isotope evidence for salt-marsh grazing in the Bronze Age Severn Estuary, UK: implications for palaeodietary analysis at coastal sites. J Archaeol Sci 35:2111–2118
Campbell G (1992) Bronze Age Plant Remains. In: Moore J, Jennings D (eds) Reading Business Park: a Bronze Age landscape. Oxford University Committee for Archaeology for the Oxford Archaeological Unit, Oxford
Challis C (2006) Iron Age and Roman features at Eastfield House, Brasenose Driftway, Oxford. Oxon 70:97–113
Chenery C, Müldner G, Evans J, Eckardt H, Lewis M (2010) Strontium and stable isotope evidence for diet and mobility in Roman Gloucester, UK. J Archaeol Sci 37:150–163
Copley MS, Berstan R, Dudd SN, Dochert G, Mukherjee AJ, Straker V, Payne S, Evershed RP (2003) Direct chemical evidence for widespread dairying in prehistoric Britain. Proc Natl Acad Sci USA 100:1524–1529
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
Fairbairn A (1999) Charred plant remains. In: Whittle A, Pollard J, Grigson C (eds) The harmony of symbols: The Windmill Hill causewayed enclosure. Oxbow Books, Oxford
Fogel ML, Tuross N (2003) Extending the limits of paleodietary studies of humans with compound specific carbon isotope analysis of amino acids. J Archaeol Sci 30:535–545
Fuller BT, Molleson TI, Harris DA, Gilmour LT, Hedges REM (2006) Isotopic evidence for breastfeeding and possible adult dietary differences from Late/Sub-Roman Britain. Am J Phys Anthropol 129(1):45–54
Grigson C (1999) The mammalian remains. In: Whittle A, Pollard J, Grigson C (eds) The harmony of symbols: The Windmill Hill causewayed enclosure. Oxbow Books, Oxford
Hamilton J, Hedges REM, Robinson M (2009) Rooting for pigfruit: pig feeding in Neolithic and Iron Age Britain compared. Antiq 83:998–1011
Hamilton-Dyer S (1997) The animal bones. In: Fulford M, Rippon S, Ford S, Timby J, Williams B. Silchester (eds) Excavations at the North Gate, on the North Walls and in the Northern Suburbs 1988 and 1991–1993, vol 28. Britannia, pp. 87–168
Hedges REM, Reynard LM (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Hedges REM, Stevens RE, Pearson JA (2006) Carbon and nitrogen stable isotope compositions of animal and human bone from Ascott-under-Wychwood long barrow. In: Benson D, Whittle A (eds) Building memories: the Neolithic Cotswold long barrow at Ascott-under-Wychwood, Oxfordshire. Oxbow, Oxford
Hedges REM, Clement JG, Thomas CDL, O’Connell TC (2007) Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. Am J Phys Anthropol 133:808–816
Hedges REM, Saville A, O’Connell TC (2008) Characterizing the diet of individuals at the Neolithic chambered tomb of Hazleton North, Gloucestershire, England, using stable isotopic analysis. Archaeometry 50:114–128
Helbaek H (1952) Early crops in southern Britain. P Prehis Soc 18:194–233
Hillman G (1981) Crop husbandry: evidence from macroscopic plant remains. In: Simmons IG, Tooley MJ (eds) The environment in British prehistory. Duckworth, London
Hoefs J (1997) Stable isotope geochemistry, 4th edn. Springer, Berlin
Jay M (2008) Iron Age diet at Glastonbury lake village: The isotopic evidence for negligible aquatic resource consumption. Oxf J Archaeol 27:201–216
Jay M, Richards MP (2006) Diet in the Iron Age cemetery population at Wetwang Slack, East Yorkshire, UK: Carbon and nitrogen stable isotope evidence. J Archaeol Sci 33:653–662
Jay M, Richards MP (2007a) The Beaker People Project: progress and prospects for the carbon, nitrogen and sulphur isotopic analysis of collagen. In: Larsson M, Parker Pearson M (Eds) From Stonehenge to the Baltic: living with cultural diversity in the third millennium BC. Archaeopress, BAR International 1692, Oxford, pp 77–82
Jay M, Richards MP (2007b) British Iron Age diet: stable isotopes and other evidence. P Prehis Soc 73:169–190
Jim S, Jones V, Ambrose SH, Evershed RP (2006) Quantifying dietary macronutrient sources of carbon for bone collagen biosynthesis using natural abundance stable carbon isotope analysis. Br J Nutr 95:1055–1062
Jones MK (1975) Seed report. In: Parrington M, Balkwill C (eds) Excavations at Broad Street. Oxoniensia 40: 5–58
Jones MK (1978) The plant remains. In: Parrington M (ed) The excavation of an Iron Age settlement, Bronze Age ring ditches and Roman features at Ashville Trading Estate, Abingdon (Oxfordshire). Oxford Archaeological Unit/CBA, Oxford/London, pp 1974–1976
Jones MK (1984) Regional patterns in crop production. In: Cunliffe BW, Miles D (eds) Aspects of the Iron Age in Central Southern Britain. Oxford University Committee for Archaeology, Oxford
Katzenberg MA, Weber A (1999) Stable isotope ecology and palaeodiet in the Lake Baikal region of Siberia. J Archaeol Sci 26:651–659
Lambrick G (1992) The development of Late Prehistoric and Roman farming on the Thames Gravels. In: Fulford M, Nichols E (eds) Developing landscapes of lowland Britain: the archaeology of the British river gravels: a review. Society of Antiquaries, London
Letts J (1993) Charred plant remains. In: Mudd A (ed) Excavations at Whitehouse Road, Oxford, 1992. Oxoniensia 58: 33–85
Levitan B (1992) Vertebrate remains. In: Moore J, Jennings D (eds) Reading Business Park: a Bronze Age landscape. Oxford University Committee for Archaeology for the Oxford Archaeological Unit, Oxford
Lewis J, Batt A (2006) The emergence of an agricultural landscape from the early-middle Bronze Age to the end of the Early Iron Age (c 1700-400BC). In: Framework Archaeology. Landscape evolution in the Middle Thames Valley: Heathrow Terminal 5 Excavations. Framework Archaeology, Oxford
Lightfoot E, Stevens RE (2012) Stable isotope investigations of charred barley (Hordeum vulgare) and wheat (Triticum spelta) grains from Danebury Hillfort: Implications for palaeodietary reconstructions. J Archaeol Sci 39:656–662
Lightfoot E, O’Connell TC, Stevens RE, Hamilton J, Hey G, Hedges REM (2009) An investigation into diet at the site of Yarnton, Oxfordshire, using stable carbon and nitrogen isotopes. Oxf J Archaeol 28:301–322
Lightfoot E, Boneva B, Miracle PT, Šlaus M, O’Connell TC (2011) Exploring the Mesolithic and Neolithic transition in the Adriatic through isotopic investigations. Antiq 85(327):73–86
Moffett L (2004) The evidence for crop-processing products from the Iron Age and Romano-British periods and some earlier prehistoric plant remains. In: Lambrick G, Allen T, Gravelly G (eds) Stanton Harcourt Oxfordshire: the development of a Prehistoric and Romano-British Community. Oxford Archaeology, Oxford
Moffett L, Robinson MA, Straker V (1989) Cereals, fruit and nuts: charred plant remains from Neolithic sites in England and Wales and the Neolithic economy. In: Milles A, Williams D, Gardner N (eds) The beginnings of agriculture. Archaeopress, Oxford, pp 243–261
Müldner G, Richards MP (2007) Stable isotope evidence for 1500 years of human diet at the city of York, UK. Am J Phys Anthropol 133:682–697
Mulville J, Grigson C (2007) The animal bones. In: Benson D, Whittle A (eds) Building memories: the Neolithic Cotswold Long Barrow at Ascott-under-Wychwood, Oxfordshire. Oxbow, Oxford
Mulville J, Levitan B (2004) The animal bone. In: Lambrick G, Allen T, Gravelly G (eds) Stanton Harcourt Oxfordshire: the development of a Prehistoric and Romano-British community. Oxford Archaeology, Oxford
Mulville J, Powell A, (2005) Iron Age animal bones. In: Lock G, Gosden C. Daly P. (eds) Segsbury Camp: excavations in 1996 and 1997 at an Iron Age hillfort on the Oxfordshire Ridgeway. University of Oxford School of Archaeology Monograph 61, Oxford
O’Connell TC, Kimball LM (2006) Stable carbon and nitrogen isotope analysis of faunal remains from the Lismore Landscape Project: A preliminary report. Unpublished report for Historic Scotland, McDonald Institute for Archaeological Research, Cambridge University, Cambridge
Parker Pearson M, Chamberlain A, Craig OE, Marshall P, Mulville J, Smith H, Chenery C, Collins M, Cook G, Craig G, Evans J, Hiller J, Montgomery J, Schwenninger J, Taylor G, Wess T (2005) Evidence for mummification in Bronze Age Britain. Antiquity 79:529–546
Parker AG, Lucas AS, Walden J, Goudie AS, Robinson MA, Allen TG (2008) Late Holocene geoarchaeological investigation of the Middle Thames floodplain at Dorney, Buckinghamshire, UK: an evaluation of the Bronze Age, Iron Age, Roman and Saxon landscapes. Geomorphol 101:471–483
Privat KL, O’Connell TC, Richards MP (2002) Stable isotope analysis of human and faunal remains from the Anglo-Saxon cemetery at Berinsfield, Oxfordshire: dietary and social implications. J Archaeol Sci 29:779–790
Redfern RC, Hamlin C, Athfield NB (2010) Temporal changes in diet: a stable isotope analysis of late Iron Age and Roman Dorset, Britain. J Archaeol Sci 37:1149–1160
Richards MP (2008) Hambledon Hill stable isotope values. In: Mercer R, Healy F (eds) Hambledon Hill, Dorset, England: excavation and Survey of a Neolithic monument complex and its surrounding landscape. English Heritage, Swindon
Richards MP, Hedges REM (1999) A Neolithic revolution? New evidence of diet in the British Neolithic. Antiquity 73:891–897
Richards MP, Hedges REM, Molleso TI, Vogel JC (1998) Stable isotope analysis reveals variations in human diet at the Poundbury Camp cemetery site. J Archaeol Sci 25:1247–1252
Robertson-Mackay R (1987) The Neolithic causewayed enclosure at Staines, Surrey: excavations 1961-3. P Prehis Soc 53:23–128
Schoeller DA (1999) Isotope fractionation: why aren’t we what we eat? J Archaeol Sci 26:667–673
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383
Serjeantson D (1996) The animal bones. In: Needham S, Spence T (eds) Refuse and disposal at Area 16 East, Runnymede. British Museum, London, pp 194–223
Smith K (1977) The excavation of Winklebury Camp, Baisingstoke, Hampshire. Proceedings of the Prehistoric Society 43:31–130
Stenhouse MJ, Baxter MS (1979) The uptake of bomb 14 C in humans. In: Berger R, Suess HE (eds) Radiocarbon dating. University of California Press, Berkeley, pp 324–341
Stevens RE, Hedges REM (2004) Carbon and nitrogen stable isotope analysis of northwest European horse bone and tooth collagen, 40,000 BP–present: palaeoclimatic interpretations. Quat Sci Rev 23:977–991
Stevens RE, Jacobi R, Street M, Germonpre M, Conard NJ, Munzel SC, Hedges REM (2008) Nitrogen isotope analyses of reindeer (Rangifer tarandus), 45,000 BP to 900 BP: Palaeoenvironmental reconstructions. Palaeogeogr Palaeoclimatol Palaeoecol 262:32–45
Stevens RE, Lightfoot E, Hamilton J, Cunliffe B, Hedges REM (2010) Stable isotope investigations of the Danebury hillfort pit burials. Oxf J Archaeol 29:407–428
Straker V (1990) Carbonised plant macrofossils. In: Saville A (ed) Hazleton North: the Excavation of a Neolithic Long Cairn of the Cotswold-Severn Group. English Heritage, London
Tieszen LL, Fagre T (1993) Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G (eds) Prehistoric human bone—Archaeology at the molecular level. Springer, Berlin, pp 121–155
Towers J, Jay M, Mainland I, Nehlich O, Montgomery J (2011) A calf for all seasons? The potential of stable isotope analysis to investigate prehistoric husbandry practices. J Archaeol Sci 38:1858–1868
Vogel JC, van der Merwe NJ (1977) Isotopic evidence for early maize cultivation in New York State. Am Antiq 42:238–242
Acknowledgements
We would like to thank Bethan Charles, Emma Evans, Jennifer Tripp and Paul Harvey for help with selecting and sampling bones. Peter Ditchfield is thanked for help with mass spectrometry. Tarmac Ltd is thanked for giving permission for the reproduction of Fig. 2. The anonymous reviewers are thanked for their comments on an earlier version of this manuscript. EL would like to thank Darwin College for financial support; RS would like to thank the Royal Society for financial support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(XLSX 22 kb)
Rights and permissions
About this article
Cite this article
Stevens, R.E., Lightfoot, E., Allen, T. et al. Palaeodiet at Eton College Rowing Course, Buckinghamshire: isotopic changes in human diet in the Neolithic, Bronze Age, Iron Age and Roman periods throughout the British Isles. Archaeol Anthropol Sci 4, 167–184 (2012). https://doi.org/10.1007/s12520-012-0089-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-012-0089-0